Что такое биосинтез белка в клетке
Содержание:
- Время существования мРНК в клетке
- Вирусы и ретротранспозоны
- Типы мРНК
- Нарушение процессинга РНК и болезни
- Перевод мРНК
- викторина
- Механизм транскрипции
- Типы РНК
- Эндогенные ингибиторы
- Обратная транскрипция
- Общая характеристика
- Транскрипция — первый этап биосинтеза белка
- Фабрики транскрипции
- 3.3. Биосинтез белка
- История исследования вопроса
- Синтез
Время существования мРНК в клетке
Матричная РНК живет в клетке очень недолго. Период существования одной молекулы характеризуется двумя параметрами:
- Функциональное время полужизни определяется способностью мРНК служить в качестве матрицы и измеряется по уменьшению количества белка, синтезируемого с одной молекулы. У прокариот этот показатель составляет примерно 2 минуты. За этот период количество синтезируемого белка уменьшается в два раза.
- Химическое время полужизни определяется уменьшением молекул информационных РНК, способных к гибридизации (комплементарному соединению) с ДНК, что характеризует целостность первичной структуры.
Химическое время полужизни обычно длиннее функционального, поскольку незначительная начальная деградация молекулы (например, единичный разрыв в рибонуклеотидной цепи) еще не препятствует гибридизации с ДНК, но уже препятствует белковому синтезу.
Время полужизни является статистическим понятием, поэтому существование конкретной молекулы РНК может оказаться значительно выше или ниже этого значения. В результате некоторые мРНК успевают транслироваться несколько раз, а другие деградируют раньше, чем закончится синтез одной белковой молекулы.
В плане деградации эукариотические иРНК гораздо стабильнее прокариотических (период полужизни составляет около 6 часов). По этой причине их гораздо легче выделять из клетки в неповрежденном виде.
Вирусы и ретротранспозоны
Некоторые вирусы также используют кДНК для превращения своей вирусной РНК в мРНК (вирусная РНК → кДНК → мРНК). МРНК используется для того, чтобы вирусные белки захватили хозяйскую клетку.
Пример этого первого шага от вирусной ДНК к кДНК можно увидеть в цикле ВИЧ-инфекции. Здесь мембрана клетки-хозяина прикрепляется к липидной оболочке вируса, что позволяет вирусному капсиду с двумя копиями РНК вирусного генома проникать в хозяина. Копия кДНК затем создается посредством обратной транскрипции вирусной РНК, процессу, облегчаемому шапероном CypA и связанной с вирусным капсидом обратной транскриптазы.
кДНК также генерируется ретротранспозонами в геномах эукариот. Ретротранспозоны — это мобильные генетические элементы, которые перемещаются внутри генома, а иногда и между геномами через промежуточные соединения РНК. Этот механизм характерен и для вирусов, за исключением образования инфекционных частиц.
Типы мРНК
Пре-мРНК и hnRNA
Предшественник мРНК (пре-мРНК) является основным транскриптом эукариотической мРНК, поскольку он выходит из матрицы ДНК. Пре-мРНК является частью группы РНК, называемой гетерогенной ядерной РНК (hnRNA). hnRNA относится ко всем одноцепочечным РНК, расположенным внутри ядра клетки, где происходит транскрипция (ДНК-> РНК), и пре-мРНК образуют большую часть этих рибонуклеиновых кислот. Пре-мРНК содержит последовательности, которые необходимо удалить или «сплайсировать» перед трансляцией в белок. Эти последовательности могут быть удалены либо посредством каталитической активности самой РНК, либо посредством действия мультибелковой структуры, называемой сплайсосомой. После этого этапа обработки пре-мРНК рассматривается как транскрипт зрелой мРНК.
Диаграмма ниже описывает структуру пре-мРНК. Пре-мРНК включает интроны и может включать или не включать 5 ‘колпачок и полиаденилированный 3’ хвост:

Моноцистронная мРНК
Моноцистронная мРНК молекула содержит последовательности экзонов, кодирующих один белок. Большинство эукариотических мРНК являются моноцистронными.
Поликистронная мРНК
Молекула полицистронной мРНК содержит экзон-кодирующие последовательности для нескольких белков. Большая часть мРНК бактерии и бактериофаги (вирусы, которые живут в бактериальных хозяевах) являются поликистронными.
Нарушение процессинга РНК и болезни
Нормальное функционирование клеток зависит от строгого контроля уровня экспрессии как РНК, кодирующих белки, так и некодирующих РНК. Такие РНК принимают участие в транскрипции, процессинга и трансляции, поддержании длины теломер и многих других событиях в клетке. Поскольку процессинг РНК включает в себя созревания молекулы РНК от той формы, закодирована в молекуле ДНК, к зрелой функциональной РНК, то нарушение этого процесса может вызвать заболевание.
Так при возникновении изоформ мРНК — например, в результате мутаций, которые приводят к активации другого сайта сплайсинга — белки, которые считываются из таких матриц могут иметь другой аминокислотный состав или быть конформационно нестабильными, что приводит к неспособности белка выполнять свои функции. Примеров альтернативного сплайсинга, который приводит к заболеваниям, множество. Так при атаксии телеангиэктазии (синдром Луи-Бар), нейродегенеративных заболеваниях со склонностью к злокачественным новообразованиям, делеция 4 нуклеотидов в 20-м интроне гена ATM (англ. Ataxia-telengiectasia mutated) приводит к активации альтернативного сплайсинга, и вызывает развитие заболевания.
РНК существуют в клетках в связанном с белками состоянии, в виде так называемых рибонуклеопротеинових комплексов (РНП, англ. RNP), состоящих из одной или более молекул РНК и зачастую многих РНК-связывающих белков (англ. RNA-binding proteins, RBP, RNABP). Собственно, выполнение соответствующими РНК своих функций происходит в таких рибонуклеопротеинових комплексах, и их нормальная активность зависит от четкого расположения белковых структур относительно третичной структуры РНК. Сбои при процессинга адекватность некодирующих РНК, так и мРНК, кодирующих эти белки, могут привести к нарушению образования этих комплексов. Например, РНК-связывающие белки, в нормальных условиях участвуют в регуляции сплайсинга, формируют нетипичные агрегаты при болезни Паркенсона и при амиотрофический боковой склерозе.
Интересным случаем является синонимичные мутации — это такие мутации в гене, приходящихся на кодирующую участок РНК и не приводят к изменению аминокислоты, они кодируют. Например, ГГТ, ГГА и ГГГ кодируют одну аминокислоту — глицин. При точечной мутации гена в третьем положении этого кодона ГГ_ (например ГГ А → ГГ Ц), аминокислота кодируемый такой мРНК не изменится — это все равно будет глицин, отсюда и название мутации — синонимична, ведь в данном случае А синонимический Ц. Долгое время считалось, что синонимичные мутации не приводят к каким-либо влияния на функционирование клетки. Однако в некоторых случаях до 25% таких синонимических мутаций могут влиять на взаимодействие со сплайсосома и приводить к альтернативному сплайсинга.
Перевод мРНК
мРНК может транслироваться на свободных рибосомах в цитоплазме с помощью молекул переноса РНК (тРНК) и множества белков, называемых факторами инициации, удлинения и терминации. Белки, которые синтезируются на свободных рибосомах в цитоплазме, часто используются клеткой в самой цитоплазме или предназначены для использования внутри внутриклеточных органелл. Альтернативно, белки, которые должны секретироваться, начинают транслироваться в цитоплазме, но как только первые несколько остатков транслируются, специфические белки транспортируют весь механизм трансляции в мембрану эндоплазматическая сеть (ЭР). Первые несколько аминокислот внедряются в мембрану ER, а остальная часть белка высвобождается во внутреннее пространство ER. Короткая последовательность удаляется из белков, которые должны секретироваться из клетки, тогда как те, которые предназначены для внутренних мембран, сохраняют этот короткий участок, обеспечивая мембранный якорь.
Более 200 заболеваний связаны с дефектами в процессинг пре-мРНК в мРНК. Мутации в ДНК или механизме сплайсинга в основном влияют на точность сплайсинга пре-мРНК. Например, аномальная последовательность ДНК может устранять, ослаблять или активировать скрытые сайты сплайсинга в пре-мРНК. Аналогично, если механизм сплайсинга не работает должным образом, сплайсосома может неправильно разрезать пре-мРНК независимо от последовательности. Эти мутации приводят к процессингу pre-mMRA в мРНК, которые будут кодировать неисправные белки. Сами аномальные мРНК также иногда являются мишенями для нонсенс-опосредованного распада мРНК, а также ко-транскрипционной деградации зарождающихся пре-мРНК. Клетки, полученные от пациентов с различными заболеваниями, включая прогерию, рак молочной железы и муковисцидоз, имеют дефекты сплайсинга РНК, причем наиболее распространенными являются рак и невропатологические заболевания.
- рибосома – Рибосомы – это ферменты, состоящие из многих белков, которые катализируют синтез белков из мРНК в процессе трансляции. Рибосомы свободно существуют в цитоплазме клетки или остаются прикрепленными к эндоплазматической сети.
- RNAP II – РНК-полимераза II – это фермент, состоящий из многих белков, который читает ДНК и синтезирует РНК в ядре клетки в процессе, называемом транскрипцией.
- транскрипция – Транскрипция – это синтез РНК из ДНК с помощью РНК-полимеразы.
- Перевод – Трансляция – это синтез белков из мРНК с участием рибосом и других белков.
викторина
1. Молекулы зрелой мРНК короткие, одноцепочечные и содержат следующие компоненты:A. аденин, цитозин, гуанин и урацил, экзоны, 5′-колпачок и 3′-полихвостB. аденин, цитозин, гуанин и урацил, интроны, экзоны, 5′-колпачок и 3′-полихвостC. аденин, цитозин, гуанин и урацил, интроныD. интроны, 5′-кепка и 3′-поли-хвост
Ответ на вопрос № 1
верно. Молекула мРНК представляет собой короткую одноцепочечную молекулу, содержащую аденин, цитозин, гуанин и урацил, экзоны, 5′-колпачок и 3′-полихвост. Интроны были сплайсированы автоматически самой мРНК или сплайсосомой.
2. Назовите местоположение и клеточный механизм, участвующий в транскрипции и трансляции мРНК.A. Транскрипция происходит в ядре под действием рибосом; трансляция происходит в цитоплазме через RNAP II.B. Транскрипция происходит в ядре под действием RNAP II; трансляция происходит в цитоплазме или на эндоплазматическом ретикулуме под действием рибосомы.C. Транскрипция происходит на клеточная мембрана благодаря действиям RNAP II; трансляция происходит в цитоплазме под действием факторов трансляции.D. Ничто из вышеперечисленного не является правильным.
Ответ на вопрос № 2
В верно. Молекула мРНК транскрибируется в ядре ферментом RNAP II и транслируется рибосомой, которая находится в цитоплазме или эндоплазматической сети клетки.
3. Какие из следующих утверждений верны в отношении различий между эукариотической и прокариотической мРНК?A. В отличие от эукариот, которые транскрибируют в ядре и транслируют в цитоплазме, прокариоты транскрибируют и транслируют мРНК одновременно в цитоплазме.B. Прокариот мРНК является преимущественно полицистронной, а эукариотическая мРНК – преимущественно моноцистронной.C. Бактериальные мРНК являются короткоживущими для обеспечения гибкости в быстро меняющихся условиях, в то время как эукариотические мРНК стабильны в течение нескольких дней.D. Все вышеперечисленное верно.
Ответ на вопрос № 3
D верно. Все вышеприведенные утверждения верны.
4
Что происходит во время важного этапа процессинга пре-мРНК в мРНК?A. Некодирующие интроны удаляются или «сращиваются».B
МРНК переводится в белок.C. Пре-мРНК экспортируется из ядра.D. Все вышеперечисленное.
Ответ на вопрос № 4
верно. Некодирующие интроны удаляются из пре-мРНК в ядре. После обработки мРНК с присоединенным 5′-колпачком и 3′-поли-А хвостом экспортируется через ядерные поры и доставляется в рибосомы, где происходит трансляция.
Механизм транскрипции
Синтез матричной РНК основан на принципе комплементарности нуклеиновых кислот и осуществляется РНК-полимеразами, которые катализируют замыкание фосфодиэфирной связи между рибонуклеозидтрифосфатами.
У прокариот мРНК синтезируется тем же ферментом, что и другие виды рибонуклеотидов, а у эукариот — РНК-полимеразой II.

Транскрипция включает 3 стадии: инициацию, элонгацию и терминацию. На пером этапе полимераза присоединяется к промотору — специализированному участку, предваряющему кодирующую последовательность. На стадии элонгации фермент наращивает цепь РНК, присоединяя к цепи нуклеотиды, комплементарно взаимодействующие с матричной цепью ДНК.
Типы РНК
В зависимости от функций, выполняемых в организме, принято выделять несколько типов рибонуклеиновой кислоты. Каждый из них имеет своё специальное обозначение.
Различные типы этого вещества и соответствующие функции РНК для наглядности можно представить в виде таблицы:
| Название | Условное обозначение | Особенности |
| Информационная (матричная) | иРНК (мРНК) | Из всей рибонуклеиновой кислоты, содержащейся в клетке, она составляет около 5%. Содержит и передаёт информацию о первичной структуре белка. Созревая, становится матрицей для синтеза полипептидной белковой молекулы. Молекулы информационной РНК присутствуют в клетке до тех пор, пока синтезируется необходимая белковая молекула. После того как матрица становится не нужна, клетка ее разрушает. |
| Рибосомальная | рРНК | Синтез рибосомальной РНК осуществляется в ядрышке. Её молекулы имеют довольно крупные габариты, состоят из из большого количества нуклеотидов — от 3000 до 5000. Составляя 80−85% всей РНК клетки, имеет несколько разновидностей, которые входят в состав рибосом, отличаясь друг от друга длиной цепи, выполняемыми функциями, а также вторичной и третичной структурой. Молекулы рибосомальной РНК считывают информацию, закодированную информационной молекулой и способствуют образованию связей между аминокислотами в белковой цепи. |
| Транспортная | тРНК | Эта разновидность рибонуклеиновой кислоты синтезируется в ядре клетки на основе матрицы ДНК, после чего выходит в цитоплазму. Характерной чертой транспортной РНК является небольшой по меркам полимерных веществ размер молекулы (по сравнению с молекулами того же вещества, которым присущи другие функции). Она может содержать около 80 мономеров. Функция этого вещества: транспорт аминокислот, являющихся строительными материалами для протеинов к месту сборки белковой молекулы. Если представить пространственную структуру молекулы нуклеиновой кислоты в виде фигуры, напоминающей листок клевера, то транспортируемая аминокислота присоединяется к его черешку. Молекула транспортной рибонуклеиновой кислоты неуниверсальна: для доставки к рибосоме каждого вида аминокислот необходима своя разновидность транспортной РНК. Всего таких видов известно около 60. |
Указанные в таблице типы РНК являются основными. Кроме них существуют и другие разновидности этого вещества. Все они в совокупности составляют единую систему, значение которой крайне велико: она направлена на считывание и воспроизведение наследственной информации через синтез белковых структур.
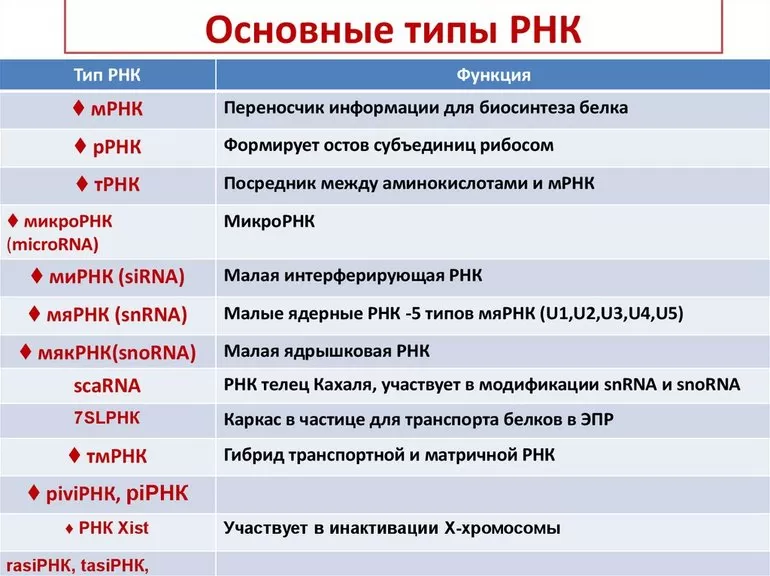
Существует ещё одна классификации РНК; согласно ей, выделяют следующие разновидности:

- Ядерная. Рапространение — ядро эукариотических клеток. Молекула собирается полимеразой 2 или 3 типов. После сборки выходит в цитоплазму клетки, где происходит созревание; потом возвращается в ядро. Участвует в процессе созревания матричной РНК. В цепи такой нуклеиновой кислоты находится много уридиновых нуклеотидов. Имеется и малый (ядрышковый) подтип.
- Цитоплазматическая. Находится под влиянием ядерной разновидности нуклеиновой кислоты. Функция — участие в антителообразовании в зрелых плазматических клетках.
- Митохондриальная. В отличие от ядерной, располагается в митохондриях.
- Пластидная. Кодирует гены, обеспечивающие процессы транскрипции и трансляции.
Эндогенные ингибиторы
У позвоночных большинство генов содержат с многочисленными сайтами CpG . Когда многие из промоторных сайтов CpG гена метилированы, ген становится подавленным (заглушенным). Колоректальный рак обычно имеет от 3 до 6 мутаций и от 33 до 66 мутаций автостопщика или пассажира. Однако подавление транскрипции (молчание) может иметь большее значение, чем мутации, в развитии рака. Например, при колоректальном раке от 600 до 800 генов транскрипционно ингибируются метилированием CpG-островков (см. Регуляцию транскрипции при раке ). Репрессия транскрипции при раке также может происходить за счет других эпигенетических механизмов, таких как измененная экспрессия . При раке груди репрессия транскрипции BRCA1 может происходить чаще за счет сверхэкспрессии микроРНК-182, чем за счет гиперметилирования промотора BRCA1 (см. ).
Обратная транскрипция
![]()
Схема обратной транскрипции
Некоторые вирусы (например, ВИЧ , вызывающий СПИД ) обладают способностью транскрибировать РНК в ДНК. У ВИЧ есть геном РНК, который обратно транскрибируется в ДНК. Полученная ДНК может быть объединена с геномом ДНК клетки-хозяина. Основной фермент, ответственный за синтез ДНК из матрицы РНК, называется обратной транскриптазой .
В случае ВИЧ обратная транскриптаза отвечает за синтез комплементарной цепи ДНК (кДНК) геному вирусной РНК. Затем фермент рибонуклеаза H переваривает цепь РНК, а обратная транскриптаза синтезирует комплементарную цепь ДНК с образованием двойной спиральной структуры ДНК («кДНК»). КДНК интегрируется в геном клетки-хозяина с помощью фермента интегразы , который заставляет клетку-хозяина генерировать вирусные белки, которые собираются в новые вирусные частицы. В ВИЧ, после этого, клетка — хозяин подвергается запрограммированной гибели клеток или апоптоз в Т — клетках . Однако в других ретровирусах клетка-хозяин остается нетронутой, поскольку вирус выходит из клетки.
Некоторые эукариотические клетки содержат фермент с активностью обратной транскрипции, называемый теломеразой . Теломераза — это обратная транскриптаза, удлиняющая концы линейных хромосом. Теломераза несет матрицу РНК, из которой она синтезирует повторяющуюся последовательность ДНК, или «мусорную» ДНК. Эта повторяющаяся последовательность ДНК называется теломер и может рассматриваться как «крышка» для хромосомы
Это важно, потому что каждый раз, когда удваивается линейная хромосома, она укорачивается. С помощью этой «мусорной» ДНК или «крышки» на концах хромосом сокращение устраняет некоторую несущественную повторяющуюся последовательность, а не последовательность ДНК, кодирующую белок, которая находится дальше от конца хромосомы.
Теломераза часто активируется в раковых клетках, чтобы раковые клетки могли бесконечно дублировать свои геномы без потери важной последовательности ДНК, кодирующей белок. Активация теломеразы может быть частью процесса, который позволяет раковым клеткам стать бессмертными
Фактор бессмертия рака посредством удлинения теломер за счет теломеразы, как было доказано, встречается в 90% всех канцерогенных опухолей in vivo, а в оставшихся 10% используется альтернативный путь поддержания теломер, называемый ALT или альтернативное удлинение теломер.
Общая характеристика
Как и все рибонуклеиновые кислоты, матричная РНК представляет собой цепь рибонуклеотидов (аденин, гуанин, цитозин и урацил), соединенных друг с другом фосфодиэфирными связями. Чаще всего мРНК имеет только первичную структуру, но в некоторых случаях — вторичную.

В клетке присутствуют десятки тысяч видов иРНК, каждый из которых представлен 10-15 молекулами, соответствующими конкретному участку в ДНК. В иРНК записана информация о структуре одного или нескольких (у бактерий) белков. Аминокислотная последовательность представлена в виде триплетов кодирующего участка молекулы мРНК.
Транскрипция — первый этап биосинтеза белка
Транскрипция — это процесс синтеза молекулы иРНК на участке молекулы ДНК.
Транскрипция (с лат. transcription — переписывание) происходит в ядре клетки с участием ферментов, основную работу из которых осуществляет транскриптаза. В этом процессе матрицей является молекула ДНК.

Специальный фермент находит ген и раскручивает участок двойной спирали ДНК. Фермент перемещается вдоль цепи ДНК и строит цепь информационной РНК в соответствии с принципом комплементарности. По мере движения фермента растущая цепь РНК матрицы отходит от молекулы, а двойная цепь ДНК восстанавливается. Когда фермент достигает конца копирования участка, то есть доходит до участка, называемого стоп-кодоном, молекула РНК отделяется от матрицы, то есть от молекулы ДНК. Таким образом, транскрипция — это первый этап биосинтеза белка. На этом этапе происходит считывание информации путём синтеза информационной РНК.
Копировать информацию, хотя она уже содержится в молекуле ДНК, необходимо по следующим причинам: синтез белка происходит в цитоплазме, а молекула ДНК слишком большая и не может пройти через ядерные поры в цитоплазму. А маленькая копия её участка — иРНК — может транспортироваться в цитоплазму.
После транскрипции громоздкая молекула ДНК остаётся в ядре, а молекула иРНК подвергается «созреванию» — происходит процессинг иРНК. На её 5’ конец подвешивается КЭП для защиты этого конца иРНК от РНКаз — ферментов, разрушающих молекулы РНК. На 3’ конце достраивается поли(А)-хвост, который также служит для защиты молекулы. После этого проходит сплайсинг — вырезание интронов (некодирующих участков) и сшивание экзонов (информационных участков). После процессинга подготовленная молекула транспортируется из ядра в цитоплазму через ядерные поры.
Транскрипция пошагово:
- РНК полимераза садится на 3’ конец транскрибируемой цепи ДНК.
- Начинается элонгация — полимераза «скользит» по ДНК в сторону 5’ конца и строит цепь иРНК, комплементарную ДНК.
- Полимераза доходит до конца гена, «слетает» с ДНК и отпускает иРНК.
- После этого происходит процесс созревания РНК — процессинг.
Проверьте себя: помните ли вы принцип комплементарности? Молекула ДНК состоит из двух спирально закрученных цепей. Цепи в молекуле ДНК противоположно направлены. Остов цепей ДНК образован сахарофосфатными остатками, а азотистые основания одной цепи располагаются в строго определённом порядке напротив азотистых оснований другой — это и есть правило комплементарности.
Фабрики транскрипции
Активные единицы транскрипции сгруппированы в ядре в дискретных участках, называемых фабриками транскрипции или эухроматином . Такие сайты можно визуализировать, позволив задействованным полимеразам расширить свои транскрипты в помеченных предшественниках (Br-UTP или Br-U) и пометив иммуно-меченую зарождающуюся РНК. Фабрики транскрипции также можно локализовать с помощью флуоресцентной гибридизации in situ или пометить антителами, направленными против полимераз. В нуклеоплазме клетки HeLa имеется ~ 10 000 фабрик , среди которых ~ 8 000 фабрик полимеразы II и ~ 2 000 фабрик полимеразы III. Каждая фабрика полимеразы II содержит ~ 8 полимераз. Поскольку наиболее активные единицы транскрипции связаны только с одной полимеразой, каждая фабрика обычно содержит ~ 8 различных единиц транскрипции. Эти единицы могут быть связаны через промоторы и / или энхансеры, при этом петли образуют «облако» вокруг фактора.
3.3. Биосинтез белка
Биосинтез белка в организме эукариот происходит в несколько этапов.
1. Транскрипция – это процесс синтеза и-РНК на матрице ДНК. Цепи ДНК в области активного гена освобождаются от ги-стонов. Водородные связи между комплементарными азотистыми основаниями разрываются. Основной фермент транскрипции РНК-полимераза присоединяется к промотору – специальному участку ДНК. Транскрипция проходит только с одной (кодоген-ной) цепи ДНК. По мере продвижения РНК-полимеразы по кодо-генной цепи ДНК рибонуклеотиды по принципу комплементарности присоединяются к цепочке ДНК, в результате образуется незрелая про-и-РНК, содержащая как кодирующие, так и некоди-рующие нуклеотидные последовательности.
2. Затем происходит процессинг – созревание молекулы РНК. На 5-конце и-РНК формируется участок (КЭП), через который она соединяется с рибосомой. Ген, т. е. участок ДНК, кодирующий один белок, содержит как кодирующие последовательности нуклеотидов – экзоны, так и некодирующие – интроны. При про-цессинге интроны вырезаются, а экзоны сшиваются. В результате на 5-конце зрелой и-РНК находится кодон-инициатор, который первым войдет в рибосому, затем следуют кодоны, кодирующие аминокислоты полипептида, а на 3-конце – кодоны-терминато-ры, определяющие конец трансляции. Цифрами 3 и 5 обозначаются соответствующие углеродные атомы рибозы. Кодоном называется последовательность из трех нуклеотидов, кодирующая какую-либо аминокислоту – триплет. Рамка считывания нуклеиновых кислот предполагает «слова»-триплеты (кодоны), состоящие из трех «букв»-нуклеотидов.
Транскрипция и процессинг происходят в ядре клетки. Затем зрелая и-РНК через поры в мембране ядра выходит в цитоплазму, и начинается трансляция.
3. Трансляция – это процесс синтеза белка на матрице и РНК. В начале и-РНК 3-концом присоединяется к рибосоме. Т-РНК доставляют к акцепторному участку рибосомы аминокислоты, которые соединяются в полипептидную цепь в соответствии с шифрующими их кодонами. Растущая полипептидная цепь перемещается в донорный участок рибосомы, а на акцепторный участок приходит новая т-РНК с аминокислотой. Трансляция прекращается на кодонах-терминаторах. Генетический код
Это система кодирования последовательности аминокислот белка в виде определенной последовательности нуклеотидов в ДНК и РНК.
Единица генетического кода (кодон) – это триплет нуклеоти-дов в ДНК или РНК, кодирующий одну аминокислоту.
Всего генетический код включает 64 кодона, из них 61 кодирующий и 3 некодирующих (кодоны-терминаторы, свидетельствующие об окончании процесса трансляции).
Кодоны-терминаторы в и-РНК: УАА, УАГ, УГА, в ДНК: АТТ, АТЦ, АЦТ.
Начало процесса трансляции определяет кодон-инициатор (АУГ, в ДНК – ТАЦ), кодирующий аминокислоту метионин. Этот кодон первым входит в рибосому. Впоследствии метионин, если он не предусмотрен в качестве первой аминокислоты данного белка, отщепляется.
Генетический код обладает характерными свойствами.
1. Универсальность – код одинаков для всех организмов. Один и тот же триплет (кодон) в любом организме кодирует одну и ту же аминокислоту.
2. Специфичность – каждый кодон шифрует только одну аминокислоту.
3. Вырожденность – большинство аминокислот могут кодироваться несколькими кодонами. Исключение составляют 2 аминокислоты – метионин и триптофан, имеющие лишь по одному варианту кодона.
4. Между генами имеются «знаки препинания» – три специальных триплета (УАА, УАГ, УГА), каждый из которых обозначает прекращение синтеза полипептидной цепи.
5. Внутри гена «знаков препинания» нет.
| Предыдущая |
История исследования вопроса
Начало изучению нуклеиновых кислот было положено ещё в середине XIX века швейцарским учёным, обнаружившим эти вещества в клеточном ядре. Он назвал их нуклеином. Наличие этих веществ в прокариотических бактериальных клетках, не содержащих ядра, было доказано несколько позднее.
Предположение о роли РНК, которую она играет в биосинтезе белковых молекул, было сделано в 1939 году. В ходе эксперимента было продемонстрировано, что РНК, кодирующая структуру гемоглобина кролика, при введении в другую клетку заставляет её синтезировать тот же самый белок. Описанный опыт наглядно продемонстрировал роль этого вещества в живом организме. Параллельно с этим ещё одно исследование показало, что клетки, активно синтезирующие белковые вещества, содержат большее количество РНК, по сравнению с другими клеточными структурами.

Механизм синтеза самой рибонуклеиновой кислоты был открыт в середине XX века, за что в 1959 году была выдана Нобелевская премия по медицине. Ещё одна аналогичная награда в этой области была выдана в связи с расшифровкой последовательности цепи из 77 нуклеотидов транспортной РНК одного из видов дрожжевых грибков.
По мнению некоторых учёных, функция РНК процессе эволюции претерпела некоторые изменения. В частности, учёный Карл Везе в 1967 году выдвинул теорию так называемого «РНК мира». Согласно его предположениям, в прокариотических организмах эта нуклеиновая кислота выполняла следующие функции:
- Шифрование, хранение и передача информации, в частности, генетической информации клетки. Сейчас, после определённых изменений, которые произошли в ходе эволюции, эту функцию стала выполнять дезоксирибонуклеиновая кислота (ДНК).
- Участие в ряде метаболических процессов, которое проявляется в их ускорении (каталитическая активность). В сегодняшнем мире эта функция принадлежит ферментам — специализированным веществам, имеющим белковую природу.
Открытие нуклеиновых кислот и успехи в исследовании их свойств и других характеристик дали мощный толчок в развитии молекулярной биологии. С этого момента и берёт начало исследование механизмов передачи информации как внутри клеток, так и между ними. Полученные экспериментально данные объясняют в том числе и механизм наследования некоторых признаков (один из основных принципов теории эволюции — наследственность).
Синтез
РНК служит матрицей для синтеза кДНК. В клеточной жизни кДНК генерируется вирусами и ретротранспозонами для интеграции РНК в геномную ДНК- мишень . В молекулярной биологии РНК очищается из исходного материала после удаления геномной ДНК, белков и других клеточных компонентов. Затем кДНК синтезируется посредством обратной транскрипции in vitro .
Очистка РНК
РНК транскрибируется из геномной ДНК в клетках-хозяевах и экстрагируется путем сначала лизирования клеток, а затем очистки РНК с использованием широко используемых методов, таких как фенол-хлороформ, колонка с диоксидом кремния и методы экстракции РНК на основе гранул. Методы экстракции различаются в зависимости от исходного материала. Например, для извлечения РНК из растительной ткани требуются дополнительные реагенты, такие как поливинилпирролидон (ПВП), для удаления фенольных соединений, углеводов и других соединений, которые в противном случае сделают РНК непригодной для использования. Чтобы удалить ДНК и белки, для разложения используются такие ферменты, как ДНКаза и протеиназа К
Важно отметить, что целостность РНК поддерживается путем инактивации РНКаз с помощью хаотропных агентов, таких как изотиоцианат гуанидиния, додецилсульфат натрия (SDS), фенол или хлороформ. Затем общая РНК отделяется от других клеточных компонентов и осаждается спиртом
Существуют различные коммерческие наборы для простого и быстрого выделения РНК для конкретных приложений. Дополнительные методы на основе шариков могут использоваться для выделения определенных подтипов РНК (например, мРНК и микроРНК ) на основе размера или уникальных участков РНК.
Синтез первой цепи
Используя фермент обратной транскриптазы и очищенные матрицы РНК, получают одну цепь кДНК (синтез первой цепи кДНК). Обратная транскриптаза M-MLV из вируса лейкемии мышей Молони обычно используется из-за ее пониженной активности РНКазы Н, подходящей для транскрипции более длинных РНК. Обратная транскриптаза AMV из вируса миелобластоза птиц также может использоваться для матриц РНК с сильными вторичными структурами (т.е. с высокой температурой плавления). кДНК обычно генерируют из мРНК для анализов экспрессии генов, таких как RT-qPCR и RNA-seq . мРНК избирательно подвергается обратной транскрипции с использованием олиго-d Т- праймеров, которые представляют собой обратный комплемент полиаденилированного хвоста на 3′-конце всей мРНК. Оптимизированная смесь праймеров oligo-dT и случайных гексамеров увеличивает шанс получения полноразмерной кДНК при одновременном снижении смещения 5 ‘или 3’. Рибосомная РНК также может быть истощена для обогащения как мРНК, так и неполиаденилированных транскриптов, таких как некодирующая РНК .
Синтез второй цепи
Результат синтезов первой цепи, гибриды РНК-ДНК, можно обрабатывать с помощью нескольких методов синтеза второй цепи или обрабатывать непосредственно в последующих анализах. Ранний метод, известный как синтез с шпилькой, основывался на образовании шпильки на 3′-конце кДНК первой цепи для запуска синтеза второй цепи. Однако затравка носит случайный характер, и шпильочный гидролиз приводит к потере информации. В процедуре Габлера и Хоффмана используется РНКаза H E. Coli для разрыва мРНК, которая заменяется ДНК-полимеразой I E. Coli и закрывается ДНК-лигазой E. Coli . Оптимизация этой процедуры основана на низкой активности РНКазы H в M-MLV для разрыва мРНК с последующим удалением оставшейся РНК путем добавления РНКазы H после трансляции ДНК-полимеразы кДНК второй цепи. Это предотвращает потерю информации о последовательности на 5′-конце мРНК.