Из чего состоит днк
Содержание:
Нуклеиновые кислоты
Нуклеиновые кислоты являются биополимерами, которые наряду с белками играют важную роль в клетках всех живых организмов. Эти соединения ответственны за хранение, передачу и реализацию наследственной информации. Что такое нуклеотиды? Это мономеры нуклеиновых кислот.
Между частями нуклеотида возникают ковалентные химические связи, которые образуются в результате реакций конденсации. Такие реакции являются обратными гидролизу. Интересным фактом является то, что молекулы ДНК обычно не только длиннее, чем молекулы РНК, но и включают в себя две цепочки, которые соединены друг с другом при помощи водородных связей, возникающих между азотистыми основаниями.
Модифицированные азотистые основания
ДНК и РНК также содержат другие (не первичные) основания, которые были модифицированы после образования цепи нуклеиновой кислоты. В ДНК наиболее распространенным модифицированным основанием является 5-метилцитозин (m 5 C). В РНК есть много модифицированных оснований, в том числе содержащихся в нуклеозидах псевдоуридина (Ψ), дигидроуридина (D), инозина (I) и 7-метилгуанозина (m 7 G).
Гипоксантин и ксантин — два из многих оснований, созданных в результате присутствия мутагена , оба из которых — в результате дезаминирования (замены аминогруппы карбонильной группой). Гипоксантин образуется из аденина, ксантин — из гуанина, а урацил — в результате дезаминирования цитозина.
Модифицированные пуриновые азотистые основания
Это примеры модифицированного аденозина или гуанозина.
| Нуклеооснование | Гипоксантин | Ксантин | 7-метилгуанин |
| Нуклеозид | Инозин I | Ксантозин X | 7-метилгуанозин m 7 G |
Модифицированные пиримидиновые азотистые основания
Это примеры модифицированного цитозина, тимина или уридина.
| Нуклеооснование | 5,6-дигидроурацил | 5-метилцитозин | 5-гидроксиметилцитозин |
| Нуклеозид | Дигидроуридин D | 5-Метилцитидин m 5 C |
Нуклеотиды
Нуклеотиды состоят из пентозы (рибозы или дезоксирибозы), остатка фосфорной кислоты и одного из пяти азотистых оснований. Пентозы – это моносахариды с пятью атомами карбона. Азотистые основания – это производные пиримидина — цитозин, урацил, тимин или производные пурина – аденин и гуанин. Нуклеотиды являются фосфорными эфирами нуклеозидов.
Нуклеозиды – это соединения азотистых оснований с рибозой или дезоксирибозой. В природе не встречается нуклеозид, который состоял бы из тимина и рибозы.
В природе более распространены нуклеотиды, фосфорилированные по пятому углеродному атому пентозы. Рибонуклеотиды имеют название в зависимости от азотистого основания: уридиловая (УМФ), гуаниловая (ГМФ), цитидиловая (ЦМФ), адениловая (дАМФ) кислоты.
Дезоксирибонуклеотиды – дизоксиадениловая (дАМФ), дезоксигуаниловая (дГМФ), дезоксиуридиловая (дУМФ), дезоксицитидиловая (дЦМФ), дезокситимидиловая (дТМФ) кислоты.
Наиболее известным соединением является АМФ (аденозинмонофосфорная кислота), которая может присоединять еще два остатка фосфорной кислоты и образовывать АТФ–аденозинтрифосфорную кислоту. Это богатое энергией соединение. При образовании связей между остатками фосфорной кислоты в нуклеотидах накапливается много энергии. При разрыве этой связи выделяется много энергии, значительно больше, чем при разрыве обычной ковалентной связи. Такая связь называется макроэргической (от греч. макрос – большой, эргон – работа) и обозначается ~. Именно макроэргические связи используются для накапливания в нуклеотидах энергии в ходе метаболизма. В молекуле АТФ две макроэргические связи. АТФ гидролитично расщепляется до АДФ (аденозиндифосфорной кислоты) или АМФ с выделением энергии. Восстанавливается из АДФ путем присоединения остатка фосфорной кислоты, и сопровождается это аккумулированием энергии. Отщепление 1 молекулы фосфорной кислоты сопровождается выделением почти 42 кДж энергии.
Источником энергии для деятельности рибосом – синтеза белка – является также гуанозинтрифосфат.
Нуклеотиды способны соединяться в полинуклеотиды – нуклеиновые кислоты.
Регуляторная РНК
Самыми ранними известными регуляторами экспрессии генов были белки, известные как репрессоры и активаторы — регуляторы со специфическими короткими сайтами связывания в областях энхансеров рядом с регулируемыми генами. Более поздние исследования показали, что РНК также регулируют гены. Существует несколько видов РНК-зависимых процессов в эукариотах, регулирующих экспрессию генов в различных точках, таких как гены, репрессирующие РНКи, посттранскрипционно , длинные некодирующие РНК , блокирующие эпигенетические блоки хроматина , и энхансерные РНК, индуцирующие повышенную экспрессию генов. Также было показано, что бактерии и археи используют системы регуляторных РНК, такие как бактериальные малые РНК и CRISPR . Файер и Мелло были удостоены Нобелевской премии по физиологии и медицине 2006 года за открытие микроРНК (миРНК), специфических коротких молекул РНК, которые могут образовывать пары оснований с мРНК.
РНК-интерференция миРНК
Уровни посттранскрипционной экспрессии многих генов можно контролировать с помощью РНК-интерференции , при которой миРНК , специфические короткие молекулы РНК, соединяются с участками мРНК и нацелены на них для деградации. Этот основанный на антисмысловой основе процесс включает этапы, на которых сначала обрабатывается РНК, чтобы она могла образовывать пару оснований с областью ее мРНК-мишени. Как только происходит спаривание оснований, другие белки направляют мРНК на разрушение нуклеазами .
Длинные некодирующие РНК
Затем с регуляцией были связаны Xist и другие длинные некодирующие РНК, связанные с инактивацией Х-хромосомы . Их роль, поначалу загадочная, как было показано Джинни Т. Ли и другими , заключалась в подавлении блоков хроматина за счет рекрутирования комплекса Polycomb, так что информационная РНК не могла быть транскрибирована с них. Дополнительные днРНК, в настоящее время определяемые как РНК из более чем 200 пар оснований, которые, по-видимому, не обладают кодирующим потенциалом, были обнаружены связанными с регуляцией плюрипотентности стволовых клеток и клеточного деления .
Энхансерные РНК
Третья основная группа регуляторных РНК называется энхансерными РНК . В настоящее время неясно, являются ли они уникальной категорией РНК различной длины или представляют собой отдельное подмножество lncRNAs. В любом случае они транскрибируются с энхансеров , которые представляют собой известные регуляторные сайты в ДНК рядом с регулируемыми ими генами. Они активируют транскрипцию гена (ов) под контролем энхансера, с которого они транскрибируются.
Регуляторная РНК у прокариот
Сначала считалось, что регуляторная РНК является эукариотическим феноменом, частью объяснения того, почему у высших организмов было обнаружено гораздо больше транскрипции, чем предполагалось. Но как только исследователи начали искать возможные регуляторы РНК в бактериях, они обнаружили и там, что называется малая РНК (мРНК). В настоящее время в поддержку теории Мира РНК обсуждается повсеместная природа систем регуляции РНК генов . Бактериальные малые РНК обычно действуют посредством антисмыслового спаривания с мРНК, подавляя ее трансляцию, либо влияя на стабильность, либо влияя на цис-связывающую способность. Также были обнаружены рибовключатели . Это цис-действующие регуляторные последовательности РНК, действующие аллостерически . Они меняют форму, когда связывают метаболиты, так что они приобретают или теряют способность связывать хроматин для регулирования экспрессии генов.
У архей также есть системы регуляторных РНК. Система CRISPR, которая недавно использовалась для редактирования ДНК in situ , действует через регуляторные РНК у архей и бактерий, обеспечивая защиту от вирусных захватчиков.
Состав
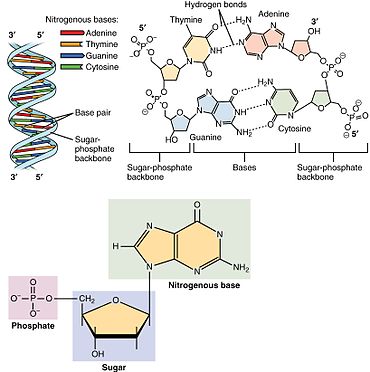
Показано расположение нуклеотидов в структуре нуклеиновых кислот: внизу слева — монофосфатный нуклеотид; его азотистое основание представляет одну сторону пары оснований. В правом верхнем углу четыре нуклеотида образуют две пары оснований: тимин и аденин (соединенные двойными водородными связями) и гуанин и цитозин (соединенные тройными водородными связями). Отдельные нуклеотидные мономеры связаны цепями на своих молекулах сахара и фосфата, образуя два «остова» ( двойную спираль ) нуклеиновой кислоты, показанные в верхнем левом углу.
Нуклео прилив состоит из трех отличительных химических субъединиц: молекулы сахара пять углерода, Нуклеиновое -The два из которых вместе называются Núcleo стороны -and одну фосфатной группы . С всеми три присоединился, нуклеотидный также называют «Nucleo бокового моно фосфат», «нуклеозид — ди — фосфат» или «нуклеозид три фосфатом», в зависимости от того , сколько фосфатов составляют группу фосфата.
В нуклеиновых кислотах нуклеотиды содержат пуриновое или пиримидиновое основание, то есть молекулу азотистого основания, также известную как азотистое основание, и называются рибонуклеотидами, если сахар представляет собой рибозу, или дезоксирибонуклеотидами, если сахар представляет собой дезоксирибозу. Отдельные молекулы фосфата повторно соединяют молекулы сахарного кольца в двух соседних нуклеотидных мономерах, таким образом соединяя нуклеотидные мономеры нуклеиновой кислоты от конца к концу в длинную цепь. Эти цепные соединения молекул сахара и фосфата создают «основу» для одинарной или двойной спирали . В любой одной цепи химическая ориентация ( направленность ) цепных соединений проходит от к ( читай : 5 простых концов к 3 простым концам), имея в виду пять углеродных сайтов на молекулы сахара в соседних нуклеотидах
В двойной спирали две нити ориентированы в противоположных направлениях, что позволяет создавать пары оснований и комплементарность между парами оснований, что важно для репликации или транскрипции кодированной информации, обнаруженной в ДНК.
Тогда нуклеиновые кислоты представляют собой полимерные макромолекулы, собранные из нуклеотидов, мономерных единиц нуклеиновых кислот . Пуриновые основания аденин и гуанин и цитозин пиримидинового основания встречаются как в ДНК, так и в РНК, в то время как пиримидиновые основания тимин (в ДНК) и урацил (в РНК) встречаются только в одном. Аденин образует пару оснований с тимином с двумя водородными связями, тогда как пары гуанина с цитозином с тремя водородными связями.
Помимо того, что они являются строительными блоками для создания полимеров нуклеиновых кислот, единичные нуклеотиды играют роль в хранении и обеспечении клеточной энергии, передаче клеточных сигналов, в качестве источника фосфатных групп, используемых для модуляции активности белков и других сигнальных молекул, а также в качестве ферментных кофакторов. , часто проводя окислительно-восстановительные реакции. Сигнальные циклические нуклеотиды образуются путем двойного связывания фосфатной группы с одной и той же молекулой сахара, соединяя 5′- и 3′- гидроксильные группы сахара. Некоторые сигнальные нуклеотиды отличаются от стандартной конфигурации однофосфатных групп тем, что имеют несколько фосфатных групп, прикрепленных к разным положениям на сахаре. Нуклеотидные кофакторы включают более широкий спектр химических групп, связанных с сахаром через гликозидную связь , включая никотинамид и флавин , и в последнем случае сахар рибозы является линейным, а не образующим кольцо, наблюдаемое в других нуклеотидах.
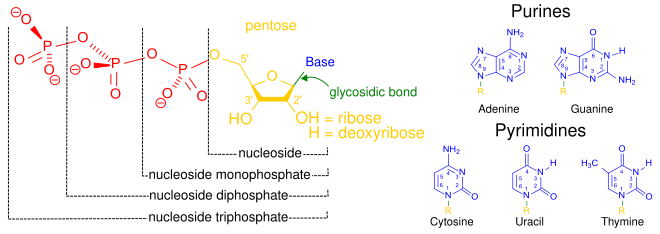
Структурные элементы три нуклео приливы -где одно-, двух- или трех-фосфаты прикреплены к ядерна стороне (в желтом, синем, зеленый) в центре: 1, нуклеотидные называют как нуклеозид моно- фосфата формируются путем добавления фосфата (в красном); Второй, добавление второго фосфата образует нуклеозид — ди — фосфат ; Третий, добавив результаты третьего фосфата в нуклеозиде три- фосфата . + Азотистое основание (азотистое основание ) обозначается как «Основание» и « гликозидная связь » (сахарная связь). Все пять основных или канонических оснований — и — показаны справа (синим цветом).
Примеры нуклеотидов, не являющихся нуклеиновыми кислотами
Наследственные болезни
Генетический код это очень многофункциональная и противоречивая структура. С одной стороны он должен хранить информацию в неизменном эталонном виде, и эта функция проявляется возможностью ДНК восстанавливать искусственные повреждения в следующем поколении. С другой же стороны, геном может быть либо поврежден, либо измениться сам, что называют мутацией.
Мутации естественное свойство генов, и последствия этих мутация бывают, как отрицательные, так и положительные. Хоть мутации и называют поломками, но это определение спорно. Некоторые мутации в чём-то ослабляют организм – именно эти мутации и ищут во время тестирования на непереносимость пищевых продуктов.
Такие мутации создают повышенные риски возникновения, какого либо заболевания при соблюдении некоторых факторов. Соответственно, если исключить эти факторы из своей жизни, то с ними будут исключены и вероятности возникновения заболевания.

Строение ДНК
ДНК – это линейный органический полимер. Его мономерные звенья – нуклеотиды, которые, в свою очередь, состоят из:
- азотистого основания;
- пятиуглеродного сахара (пентозы);
-
фосфатной группы (рисунок 1).
При этом, фосфатная группа присоединена к 5′-атому углерода моносахаридного остатка, а органическое основание — к 1′-атому.
Основания в ДНК бывают двух типов:
- Пуриновые: аденин ( А ) и гуанин (G);
-
Пиримидиновые: цитозин (С) и тимин (Т);(рисунок 2),
Строение нуклеотидов в молекуле ДНК
В ДНК моносахарид представлен 2′-дезоксирибозой, содержащей только 1 гидроксильную группу (ОН), а в РНК — рибозой, имеющей 2 гидроксильные группы (OH).
Нуклеотиды соединены друг с другом фосфодиэфирными связями, при этом фосфатная группа 5′-углеродного атома одного нуклеотида связана с З’-ОН-группой дезоксирибозы соседнего нуклеотида (рисунок 1). На одном конце полинуклеотидной цепи находится З’-ОН-группа (З’-конец), а на другом — 5′-фосфатная группа (5′-конец).
Уровни структуры ДНК
Принято выделять 3 уровня структуры ДНК:
- первичную;
- вторичную;
- третичную.
Первичная структура ДНК – это последовательность расположения нуклеотидов в полинуклеотидной цепи ДНК.
Вторичная структура ДНК стабилизируется водородными связями между комплементарными парами оснований и представляет собой двойную спираль из двух антипараллелных цепочек, закрученных вправо вокруг одной оси.
Общий виток спирали- 3,4нм, расстояние между цепочками 2нм.
Третичная структура ДНК – суперсперализация ДНК. Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы, что часто вызвано ковалентным соединением их открытых концов. Суперспиральная структура ДНК обеспечивает экономную упаковку очень длинной молекулы ДНК в хромосоме. Так, в вытянутой форме длина молекулы ДНК составляет 8 см, а в форме суперспирали укладывается в 5 нм.
Правило Чаргаффа
Правило Э. Чаргаффа – это закономерность количественного содержания азотистых оснований в молекуле ДНК:
- У ДНК молярные доли пуриновых и пиримидиновых оснований равны: А+ G = C + Т или (А + G)/(C + Т)=1.
- В ДНК количество оснований с аминогруппами (А +C) равно количеству оснований с кетогруппами (G + Т): А +C= G + Т или (А +C)/(G + Т)= 1
- Правило эквивалентности, то есть : А=Т, Г=Ц; А/Т = 1; Г/Ц=1.
- Нуклеотидный состав ДНК у организмов различных групп специфичен и характеризуется коэффициентом специфичности: (Г+Ц)/(А+Т). У высших растений и животных коэффициент специфичности меньше 1, и колеблется незначительно: от 0,54 до 0,98, у микроорганизмов он больше 1.
Самодополняемость и шпильки
Последовательность РНК, имеющая внутреннюю комплементарность, в результате чего она сворачивается в шпильку.
Само-комплементарность относится к тому факту, что последовательность ДНК или РНК может складываться сама по себе, создавая структуру, подобную двунитевой. В зависимости от того, насколько близко друг к другу находятся части последовательности, которые дополняют друг друга, нить может образовывать петли шпильки, соединения, выпуклости или внутренние петли. РНК с большей вероятностью будет образовывать такие структуры из-за связывания пар оснований, не наблюдаемого в ДНК, например связывания гуанина с урацилом.
Последовательность РНК, показывающая шпильки (крайний правый и крайний верхний левый) и внутренние петли (нижняя левая структура)
Комплементарность пар оснований ДНК и РНК
Комплементарность между двумя антипараллельными цепями ДНК. Верхняя прядь идет слева направо, а нижняя прядь идет справа налево, выравнивая их.
Слева: в нуклеотидных пар оснований , которые могут образовывать в двухцепочечной ДНК . Между A и T есть две водородные связи, а между C и G — три. Справа: две комплементарные цепи ДНК.
Комплементарность достигается за счет четких взаимодействий между азотистыми основаниями : аденином , тимином ( урацил в РНК ), гуанином и цитозином . Аденин и гуанин — пурины , а тимин, цитозин и урацил — пиримидины . Пурины крупнее пиримидинов. Оба типа молекул дополняют друг друга и могут образовывать пары оснований только с противоположным типом азотистых оснований. В нуклеиновой кислоте нуклеиновые основания удерживаются вместе водородными связями , которые эффективно работают только между аденином и тимином и между гуанином и цитозином. Комплемент оснований A = T имеет две водородные связи, в то время как пара оснований G C имеет три водородные связи. Все другие конфигурации между азотистыми основаниями препятствовали бы образованию двойной спирали. Нити ДНК ориентированы в противоположных направлениях, они считаются антипараллельными .
| Нуклеиновая кислота | Нуклеооснования | Базовый комплект |
| ДНК | аденин (A), тимин (T), гуанин (G), цитозин (C) | А = Т, G С |
| РНК | аденин (A), урацил (U), гуанин (G), цитозин (C) | А = U, G C |
Комплементарная цепь ДНК или РНК может быть сконструирована на основе комплементарности азотистых оснований. Каждая пара оснований, A = T по сравнению с G ≡ C, занимает примерно одно и то же пространство, что позволяет формировать скрученную двойную спираль ДНК без каких-либо пространственных искажений. Водородная связь между азотистыми основаниями также стабилизирует двойную спираль ДНК.
Комплементарность цепей ДНК в двойной спирали позволяет использовать одну цепочку в качестве матрицы для построения другой. Этот принцип играет важную роль в репликации ДНК , закладывая основы наследственности , объясняя, как генетическая информация может быть передана следующему поколению. Комплементарность также используется в транскрипции ДНК , которая генерирует цепь РНК из матрицы ДНК. Кроме того, вирус иммунодефицита человека , одноцепочечный РНК-вирус , кодирует РНК-зависимую ДНК-полимеразу ( обратную транскриптазу ), которая использует комплементарность для катализа репликации генома. Обратная транскриптаза может переключаться между двумя родительскими геномами РНК путем рекомбинации с выбором копии во время репликации.
Механизмы репарации ДНК, такие как контрольное считывание , основаны на комплементарности и позволяют исправлять ошибки во время репликации ДНК путем удаления несовпадающих нуклеотидных оснований. В целом, повреждения в одной нити ДНК , могут быть устранены путем удаления поврежденного участка и его замены с использованием комплементарность , чтобы скопировать информацию из другой цепи, как это имеет место в процессах ремонта рассогласования , нуклеотидной эксцизионной репарации и базовой эксцизионной репарации .
Нити нуклеиновых кислот могут также образовывать в которых одноцепочечная ДНК может легко отжигаться с комплементарной ДНК или РНК. Этот принцип лежит в основе широко используемых лабораторных методов, таких как полимеразная цепная реакция , ПЦР.
Две цепи комплементарной последовательности называются смысловой и антисмысловой . Смысловая цепь, как правило, представляет собой транскрибируемую последовательность ДНК или РНК, которая была образована при транскрипции, в то время как антисмысловая цепь представляет собой цепь, комплементарную смысловой последовательности.
Нуклеиновые кислоты.
20 ноября, 2009
Нуклеиновые кислоты (НК) – высокомолекулярные линейные гетерополимеры, мономерами которых являются мононуклеотиды, соединенные между собой 3′,5′-фосфодиэфирной связью.
Нуклеотид состоит из:
- азотистого основания (пуринового или пиримидинового),
- углеводного компонента (пентозы – рибозы или дезоксирибозы),
- остатка фосфорной кислоты (от 1 до 3).
В зависимости от типа пентозы НК подразделяются на ДНК или РНК. Название кислот «нуклеиновые»: от слова «nucleus» – ядро. НК находятся в цитозоле, ядре и митохондриях у эукариотов и в хлоро-пластах в растительных клетках.
Химический состав НК и нуклеотидов
- В условиях мягкого щелочного гидролиза НК распадаются до нуклеотидов.
- При нагревании до 145оС в водном аммиаке – до нуклеозидов.
- В результате кислотного гидролиза – до азотистых оснований и пентоз.
Общая характеристика азотистых оснований:
— по строению подразделяются на пуриновые и пиримидиновые;
— по распространению и степени встречаемости – мажорные (основные, преобладающие) и минорные (редко встречающиеся). Из пуриновых преобладают аденин (А) и гуанин (Г), из пиримидиновых – цитозин (Ц) и урацил (У) в РНК и цитозин (Ц) и тимин (Т) – в ДНК. Для всех азотистых оснований характерна лактим-лактамная таутомерия.
А) Мажорные пуриновые основания:
аденин гуанин (лактимная форма) гуанин (лактамная форма)
Б) Мажорные пиримидиновые:
урацил (лактим-лактамные формы) цитозин (лактим-лактамные формы)
тимин (лактим-лактамные формы)
В) Минорные азотистые основания. Как правило, они по строению являются производными мажорных или могут быть нетипичными для данной кислоты. Например, в РНК обычно нет тимина, он входит в состав ДНК, но в одной из петель тРНК обнаруживают тимин.
Минорные азотистые основания могут быть продуктами реакций:
- восстановления (например, дигидроурацил, входит в состав D-петли тРНК),
- метилирования (метилирование может происходить у атомов, входящих:
— в циклическую структуру пурина или пиримидина (например, 7-метилгуанин),
— его заместителей (чаще по атомам водорода аминогруппы),
— реже по гидроксильным группам углеводного компонента.
нетипичного соединения (например, псевдоуридиловая кислота, в тРНК).
Биологическая роль минорных азотистых оснований:
- необходимы для формирования вторичной структуры НК (например, формирования петель в тРНК),
- выполняют защитную функцию (например, метилированные участки в мРНК),
- метилированные участки могут выполнять роль маркеров, по которым специальные регуляторные белки распознают участки повреждения в ДНК, участки начала матричных синтезов,
- метилирование используется также для распознавания вновь синтезированной ДНК и родительской (во время деления клетки и синтеза ДНК).
Углеводные компоненты НК:
b-D-рибоза в РНК b-D-дезоксирибоза в ДНК
Нумерация атомов в углеводе обозначается цифрой со штрихом (например, третий атом углерода 3′).
Соединяется углеводный компонент с пуриновыми азотистыми основаниями 9,1′-N-гликозидной связью, с пиримидиновыми – 1,1′-N-гликозидной связью.
Остаток фосфорной кислоты (1, 2 или 3) может находиться у 3′-, 5′-атома пентозы, реже у 2′-го.
Структурная организация нуклеиновых кислот
Первичная структура – определенная последовательность нуклеотидов в цепи. Образована фосфодиэфирными связями. Начало цепи – 5′-конец (на его конце фосфатный остаток), конец, завершение цепи, обозначается как 3′(ОН)-конец.
Как правило, в образовании самой цепи азотистые основания не участвуют, но водородные связи между комплементарными азотистыми основаниями играют важную роль в формировании вторичной структуры НК:
- между аденином и урацилом в РНК или аденином и тимином в ДНК образуются 2 водородные связи,
- между гуанином и цитозином – 3.
Для НК характерна линейная, а не разветвленная структура. Кроме первичной и вторичной структуры для большинства НК характерна третичная структура – например, ДНК, тРНК и рРНК.
Связывание нуклеотидов
Для создания цепей полимера (или нескольких единиц), которые приводят к образованию РНК и ДНК, нуклеотиды соединяются друг с другом через сахарофосфатный скелет, который образуется, когда фосфат одного нуклеотида присоединяется к сахару другого. Это возможно благодаря сильным ковалентным связям, называемым фосфодиэфирными связями.
Поскольку ДНК представляет собой двухцепочечную молекулу, две из этих полимерных цепей должны присоединяться друг к другу, как лестница. «Ступеньки» состоят из пар нуклеотидов, которые соединяют две стороны лестницы с помощью водородных связей. Что такое нуклеотид? Это структурная единица ДНК, которая состоит из азотистого основания и сахар-фосфатной основной цепи, состоящей из фосфатной группы и сахара. ДНК состоит из многих нуклеотидов, которые содержат и защищают генетические коды организма.